One Mushroom, Two Patients: A Practitioner’s Case for Targeted Lion’s Mane Formulation

What an obscure mouse strain reveals about antigen uptake, immune reactivity, and gut barrier function.
In functional medicine, we think about PUFA (polyunsaturated fatty acid) metabolism constantly. Yet we don’t often think about where that metabolism actually happens first: in the gut itself, in intimate conversation with the microbiome. This month, Romilly Hodges, CNS, walks us through a fascinating scientific story, starting with a single gene in an obscure mouse strain that provides a surprisingly rich window into fatty acid metabolism in the gut, including how it shapes barrier function (broadening our thinking beyond tight junctions), immune reactivity, and antigen uptake. The implication is that we need to be considering the microenvironment of PUFA metabolism in the digestive tract.
This is new territory, yet in functional medicine we already have impactful and relevant tools. One of the great privileges of functional medicine practice is that we can often apply emerging science sooner than conventional medicine because our systems biology lens allows for mechanistic extrapolation and our interventions are so safe and already known to be beneficial. For patients in whom immune reactivity and inflammation are central to the picture, shifting our thinking to include the gut PUFA-microbiome axis could be powerful. Not least, microbial-PUFA interactions in the gut are an area ripe for further exploration. – DrKF
Article written by Romilly Hodges, CNS, FMCP
Key article takeaways
- Gut permeability is more than a tight-junction issue.
Emerging research suggests dietary antigens can cross the intestinal barrier through goblet cell-associated antigen passages, creating another potential pathway for immune activation beyond paracellular “leakiness.” - PUFA metabolism may shape immune reactivity in the gut.
Arachidonic acid–derived leukotrienes may influence antigen uptake and inflammatory signaling, highlighting the importance of dietary fat quality, omega-3 intake, and the broader inflammatory milieu. - The microbiome is an active partner in PUFA metabolism and gut barrier health.
Beneficial microbes and their metabolites can support mucus production, goblet cell function, and a more balanced PUFA profile, whereas dysbiosis may promote inflammatory pathways.
How DPEP1, Leukotrienes, and Dietary Antigen Uptake May Shape Gut Barrier Function
There is a strain of mouse that is naturally resistant to reacting to dietary triggers. Specifically, in this mouse’s case, it does not develop anaphylaxis when exposed to an oral food challenge with a food such as peanut or egg, even though it is sensitized to that food. It will, however, respond to peritoneal administration, leading researchers to conclude that the genetically protective trait in these mice must relate to dietary antigen uptake from the intestine. These mice are known in the lab science world as C57BL/6 (sadly, not a great name for general discussion purposes, but we’ll do our best – it leads us to some new, interesting, and practical understanding).
This mysterious, protective phenomenon recently led a group of researchers from Yale University School of Medicine and Northwestern University School of Medicine to try to identify exactly what conferred this benefit. What they found has profound implications for future treatment of patients with anaphylaxis (and was the primary context for their exploration. However, it also illuminates important new aspects of gastrointestinal barrier function and integrity that have implications for functional medicine interventions across a wide spectrum of conditions.
Let’s go deeper into the science for a moment…
Surprisingly perhaps, through genetic screening, the researchers found just one responsible gene – dipeptidase 1 (DPEP1). DPEP1 catabolizes arachidonic acid-derived eicosanoids, specifically cysteinyl leukotrienes. While these leukotrienes are already known to be responsible for the most aggressive allergic reactions, this discovery was new: leukotrienes’ role in a specific channel of food allergen transport across the intestinal membrane and the essentiality of this transport for inducing an inappropriate immune response.
Eicosanoids are bioactive lipid mediators synthesized endogenously from polyunsaturated fat (PUFA) precursors. PUFAs can be incorporated into cell phospholipid membranes as a form of storage for later use. Those synthesized from omega-6 PUFAs like arachidonic acid favor pro-inflammatory mediators, whereas those from omega-3 generally increase anti-inflammatory and pro-resolving eicosanoids. When we recommend to our patients and clients that they increase the omega-3-to-omega-6 fatty acid ratio, it’s this modulation of the body’s overall inflammatory balance that we’re primarily targeting. We haven’t, however, always thought as much about the role of these PUFAs in the digestive tract, including for barrier integrity and microbial diversity (more on microbes in a bit – they are also intricately involved in our PUFA metabolism).
Back to the study of those C57BL/6 mice and the arachidonic acid cascade. It’s worth teasing this out a little to get the full picture. In the image below, you’ll see how arachidonic acid can be metabolized into active prostaglandins (“PG”s), leukotrienes (“LT”s), and more.
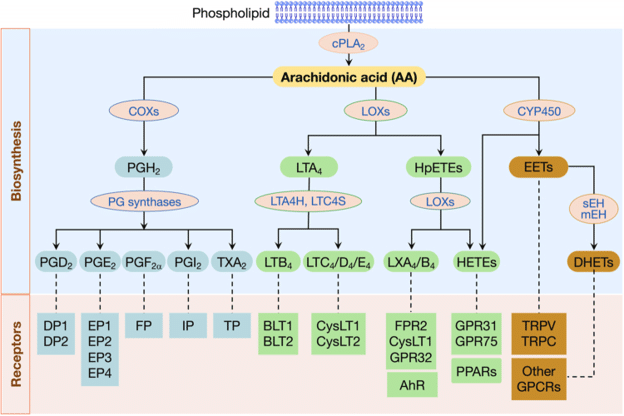
Figure 1: Biosynthesis of Eicosanoids from Arachidonic Acid. Image Source
DPEP1, which C57BL/6 mice express more of, not less, breaks down leukotriene LTD4 to LTE4 (not shown at that level in the image above). This is important because LTD4 and its cysteinyl derivatives are active in translocating antigens across the intestinal barrier into blood circulation, and mediating allergic responses. LTE4 is not. More DPEP1 breaks down LTD4 more quickly, reducing its effects. Interestingly, too, these LTD4 cysteinyl leukotrienes increase intestinal permeability to dietary antigens via goblet cells–i.e., they are not reliant on paracellular leakiness (more on this below, too).
You might be wondering how the researchers determined that this gene was so essential for inciting an allergic response? First, they tested whether oral administration of LTD4 increased antigen transport across the GI epithelium–it did. Then, conveniently, they used an existing drug (Zileuton, approved for asthma), which blocks the production of leukotrienes, to show that anaphylaxis-susceptible mice (not the C57BL/6 strain) treated with Zileuton absorbed less antigen protein into their bloodstream and showed few signs of an allergic reaction. “It was actually shocking how well Zileuton worked,” said co-senior study author, Stephanie Eisenbarth MD, PhD, “…if you had asked me six or seven years ago if leukotrienes regulate how much allergen gets in, I think almost everyone would have said no.” The researchers are now recruiting for a small human study to test the medication in allergic patients. The potential target use case is when patients are at higher risk of exposure (such as on an airplane flight or at a birthday party).
Three Functional Medicine Takeaways for Gut Barrier Health and Immune Reactivity
Of course, life-saving interventions of all kinds are hugely important for our allergy patients/clients. However, there is a lot more we can glean from these findings (even as they are pre-clinical still) when we don our functional medicine hats that applies to managing gut permeability and subsequent immune activation and reactivity. We’re not talking about acute allergic reactions now, but the much wider number of chronic conditions that involve that central gut-immune axis and the opportunity to promote a healthier intestinal barrier function.
Takeaway #1 Gut Permeability is About More Than Tight Junctions
It’s likely you have seen a representation of intestinal permeability that looks something like this:
Figure 2: Intestinal tight junction permeability (Image Source)
This kind of intestinal permeability representation is what we’re most used to seeing in functional medicine education–a breakdown in the tight junctions between epithelial cells that can allow larger molecules to enter the lamina propria than would normally be permitted. This can lead to immune dysregulation and inflammation. This is what we assume we’re testing for with the lactulose-mannitol and zonulin tests.
However, there are other ways that potential antigens (immune-activating molecules) that would normally be excluded from passage can be transported across the gut lining: M-cell mediated transcytosis in Peyer’s Patches, receptor-mediated transcytosis across enterocytes, transepithelial dendritic cells (TEDs), and goblet cell- and secretory antigen passages (GAPs and SAPs). Controlled translocation is part of the normal process of developing tolerance, whereby the body’s cells “sample” contents of the digestive tract–primarily food and microbes–to determine an appropriate response (including nonresponse). However, it too can become dysfunctional, cause sensitization to dietary or microbial components, and is thought to potentially lead to delayed-type hypersensitivity responses such as non-Celiac gluten sensitivity, depression, anxiety, joint pain, and headache. Autoimmune disease may also be related to dietary and microbial triggers via these mechanisms.
In the case of the C57BL/6 mouse study, the dysfunctional “leakiness” wasn’t related to a breakdown of tight junctions–it occurred through GAPs (goblet cell-associated antigen passages) which, under the guidance of specific leukotriene (LTD4) signals in their immediate environment, switch from a carefully controlled process of antigen sampling to one that is dysregulated. It becomes essential, then, to think of “leakiness” beyond tight junction breakdown.

Figure 3: Functions of Goblet Cells (Image Source)
Importantly, lactulose-mannitol and zonulin tests aren’t thought to measure all these types of leakiness. Tests for GAPs and other transcytosis dysfunction states are lacking.
Takeaway #2: Consider Dietary Arachidonic Acid and Inflammatory Factors in the Digestive Tract
As an astute reader, you’re probably already making the connection… leukotrienes derive from arachidonic acid. That arachidonic acid is either something you directly consumed in your last meal and is hanging around in the digestive tract, or is hanging out in your cell phospholipid membranes (from previous consumption or metabolism of other dietary omega-6 fats). This means that we can directly influence the availability of leukotriene substrate–and its metabolism–through diet and lifestyle choices.
Reduce foods that contain excess arachidonic acid
An obvious strategy for reducing excess antigen translocation in susceptible individuals is then to limit arachidonic acid-rich foods like meat, poultry, eggs, fish and dairy products. This doesn’t mean avoiding them entirely–these animal food groups still provide important nutrients (amino acids, B vitamins, zinc, iron, and betaine) that are hard to replace entirely in other foods. Vitamin B12 is only available from animal food sources or through supplementation. Arachidonic acid also actually performs essential functions when a robust immune response is needed, such as in the case of infections. But often there is room to dial down the volume on arachidonic acid intake that is present in excess.
Fat profiles in animal foods also tend to vary according to the type of feed used in animal rearing. Grass-fed cattle, for example, contain less omega-6 PUFAs (including arachidonic acid) than their grain-fed counterparts. They also tend to have more omega-3 fats. Animals fed soy protein appear to upregulate arachidonic acid production and metabolism. Labeling, however, is a bit of a minefield with various loopholes in the use of the terms “grass-fed” and “grass-finished,” which result in a wide variability in the final composition of the food. Regenerative agriculture practices, which take into account healthy soil, biodiversity, and general imitation of natural systems, may lead to optimal nutritional profiles (even as they are challenging to scale).
Optimize the omega-6:omega-3 ratio, with nuance
Omega-3 fatty acids promote a more anti-inflammatory environment, discouraging the formation of pro-inflammatory leukotrienes from arachidonic acid. Omega-3 and omega-6 fats also compete for the same elongation and desaturation enzymes, meaning that consuming more omega-3 fats can inhibit the conversion of, say, linoleic acid (an omega-6) to arachidonic acid.
Our ancestors during the Paleolithic era consumed a ratio of about 4:1 omega-6:omega-3 fats. Today’s Western diet comes in at around 20:1. Renowned NIH PUFA researcher Artemis Simopoulos had this to say about these ratios and disease outcomes:
“In the secondary prevention of cardiovascular disease, a ratio of 4/1 was associated with a 70% decrease in total mortality. A ratio of 2.5/1 reduced rectal cell proliferation in patients with colorectal cancer, whereas a ratio of 4/1 with the same amount of omega-3 PUFA had no effect. The lower omega-6/omega-3 ratio in women with breast cancer was associated with decreased risk. A ratio of 2–3/1 suppressed inflammation in patients with rheumatoid arthritis, and a ratio of 5/1 had a beneficial effect on patients with asthma, whereas a ratio of 10/1 had adverse consequences.”
Suffice to say that the pre-2025 nutritional target recommendations (omega-6: 17 g/d for males and 12 g/d for females; omega-3: 1.6 g/d for males and 1.1 g/d for females) resulting in a ratio of around 11:1 are likely to be very inadequate for protecting health. (Note that guidelines are changing rapidly at the moment). Somewhere between 4:1 to 1:1 is likely more appropriate.
A little nuance is also appropriate in our conversation about the omega-6:omega-3 ratio. These two categories cover many different types of fats, with very different effects in the body. Linoleic acid, therefore, is likely less problematic than direct arachidonic acid sources (even as linoleic can be converted to arachidonic when needed). And arachidonic acid needs certain environmental signals before it starts producing pro-inflammatory eicosanoids. Dr. Fitzgerald’s recent conversation with Dr. Bill Harris has more depth on this subject for those who want to read more. The ‘milieu’ therefore matters…
Promote an anti-inflammatory milieu
The biosynthesis of leukotrienes from arachidonic acid is not a given, even if arachidonic acid is available for use. It requires a pro-inflammatory environmental stimulus that activates phospholipase A2, LOX enzymes, and other enzymes in the pathway. Reducing pro-inflammatory stimuli, whether from diet or environmental toxins, is therefore relevant.
I already mentioned that omega-3 fatty acids promote a more anti-inflammatory environment. Plant polyphenols such as curcumin, quercetin, and olive oil phenolics, as well as kaempferol and myricetin, also inhibit phospholipase A2 and LOX enzymes. Diets high in leafy green vegetables, herbs, spices (including turmeric), teas, berries, unrefined olive oil, onions, apples, and capers provide good amounts of these phytochemicals. The epigenetically-targeted diet we studied (which is central in Dr. Fitzgerald’s Younger You program) provides ample amounts of these foods.
Takeaway #3: Consider the Role of the Microbiota in Arachidonic Acid Metabolism
It’s time to circle back to microbes. It turns out that our microbiota is highly involved in lipid and eicosanoid metabolism in the gut, and that the balance of species can lead to more or less leukotriene biosynthesis and alter the function of barrier cells.
Certain beneficial commensals actively reduce the pool of arachidonic acid available for pro-inflammatory eicosanoid synthesis. Acetate, propionate, and butyrate—short-chain fatty acids produced by the commensal microbial fermentation of dietary fiber—promote healthy goblet cell function, including maintaining a healthy mucus layer that promotes tolerogenic antigen sampling and avoids excess inflammation. Lactobacillus species can convert the omega-6 linoleic acid into a metabolite called HYA, diverting it away from the arachidonic acid pathway. Certain beneficial strains of E. coli (notably Escherichia coli Nissle 1917) produce anti-inflammatory long-chain fatty acids with their own moderating effects.
Incidentally, omega-3 fatty acids can enhance microbial diversity and can shift the microbiome towards commensals that produce SCFA, including Lactobacillus, Bifidobacterium, and Akkermansia. So can diets rich in different kinds of prebiotic (fiber and plant phytochemicals) and probiotic (live culture) foods.
A dysbiotic microbial community, by contrast, steers things the other way. In lab studies, dietary supplementation with arachidonic acid is associated with an increase in the Escherichia-Shigella genus and reduced Bifidobacterium pseudolongum. This leads to increased expression of inflammatory signaling molecules including IL-1beta. Some organisms can promote phospholipase activity, unlocking arachidonic acid from cell membrane phospholipid stores and making it available for conversion to pro-inflammatory leukotrienes and prostaglandins. There is even preliminary evidence (an abstract at the 2026 AAAAI conference, not peer-reviewed and published) from mRNA sequencing data of a bacterial leukotriene biosynthesis pathway—with homology to the human pathway—enriched in children with asthma during viral infections, along with bacterial gamma glutamyl transferase activity that converts LTC4 to the more potent LTD4. It is likely, although not yet conclusively demonstrated, that this can also happen in the intestinal lumen.
Gut Health, PUFA Metabolism, and the Functional Medicine Opportunity
It is truly remarkable how scientific advancements continue to validate the mediators we’ve long recognized and the tools we’ve long used in functional medicine, even as they aren’t yet widely recognized through the systems medicine lens. However, for functional medicine practitioners, the evolving knowledge base provides a way to deepen our understanding of the tools we use and further refine their application within the framework of interconnected systems medicine and modifiable lifestyle factors.
As always, leave your thoughts and comments below.
Related Content
Artemis Simopoulos and Dr. Fitzgerald podcast
Dr. Bill Harris and Dr. Fitzgerald podcast
Related posts





